麦克斯韦方程组
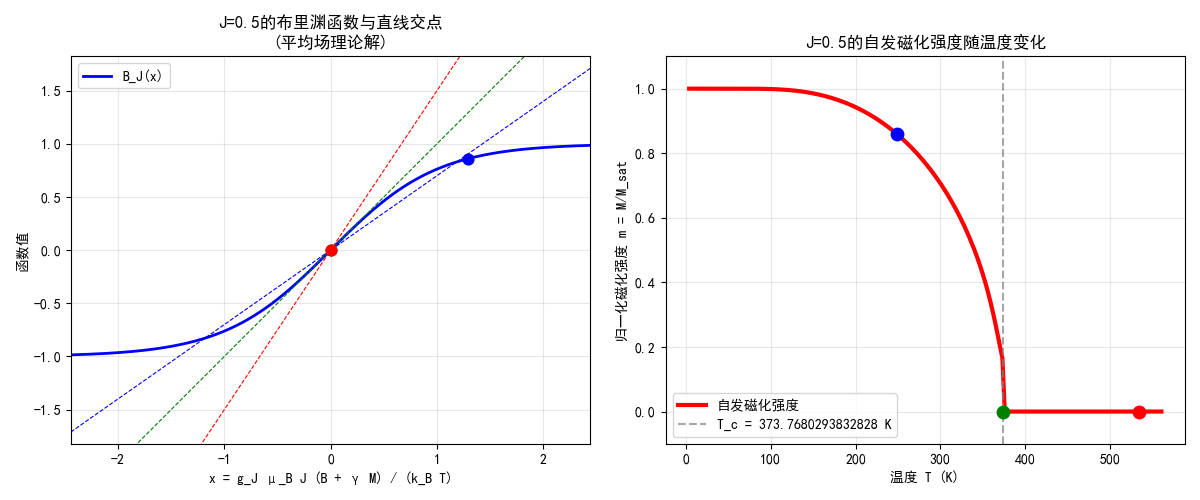
麦克斯韦方程组在静电学、静磁学和电磁感应的电磁学基础上,我们总结出了麦克斯韦方程组:
\begin{align}
\nabla\cdot\vec{E}&=\dfrac{\rho}{\varepsilon_0}\\
\nabla\cdot\vec{B}&=0\\
\nabla\times\vec{E}&=-\dfrac{\partial \vec{B}}{\partial t}\\
\nabla\times\vec{B}&=\mu_0 \vec{J}+\mu_0\varepsilon_0\dfrac{\partial \vec{E}}{\partial t}
\end{align}有介质存在时,改为:
\begin{align}
\nabla\cdot\vec{D}&=\rho_f\\
\nabla\cdot\vec{B}&=0\\
\nabla\times\vec{E}&=-\dfrac{\partial \vec{B}}{\partial t}\\
\nabla\times\vec{H}&=\vec{J}_f+\dfrac{\partial \vec{D}}{\partial t}
...
电磁感应
电磁感应磁场的变化产生电场磁通量的改变会产生电动势,分为:
感生电动势:由于磁场的变化而产生的电动势。
动生电动势:由于闭合回路磁场中运动(面积改变)而产生的电动势。
法拉第电磁感应定律指出了感应电动势的大小,而楞次定律指出了感应电动势的方向:
\mathcal{E}=-\dfrac{d\Phi_B}{dt}其中,$\Phi_B$为磁通量。楞次定律指出感应电动势的方向总是与磁通量的变化方向相反。
将其写为积分的形式:
\oint \vec{E}\cdot d\vec{l}=-\dfrac{d}{dt}\int_S \vec{B}\cdot d\vec{S}即:
\nabla\times \vec{E}=-\dfrac{\partial \vec{B}}{\partial t}电感和磁场的能量根据法拉第电磁感应定律,如果往一个线圈中通电,其产生的磁场会在另一个线圈中产生磁通量:
\Phi_2=M_{21}I_1其中,$M_{21}$为两个线圈之间的互感系数,描述两个线圈的磁场耦合程度。可以计算出:
M_{21}=\frac{\mu_0}{4\pi}\oint\oint \frac{d ...
静磁学
静磁学恒定电流定义电流为:
I=\lim_{\Delta t\to 0}\dfrac{\Delta Q}{\Delta t}=\lim_{\Delta t\to 0}\dfrac{ne\Delta V}{\Delta t}=\lim_{\Delta t\to 0}\dfrac{nevS\Delta t}{\Delta t}=nevS定义电流密度为:
J=\frac{dI}{dS_{\perp}}欧姆定律指出:
I=U/R=El S/\rho l=ES/\rho其中,电阻$R=\rho l/S$,$\rho$为电阻率。相应地,电流密度为:
\vec{J}=\sigma \vec{E}其中,$\sigma=1/\rho$为电导率。
电流满足连续性方程:
\nabla\cdot \vec{J}=-\dfrac{\partial \rho}{\partial t}若电流是恒定的,即$\rho$不随时间变化,则有:
\nabla\cdot \vec{J}=0静磁学的核心是恒定电流,尽管电流是非静止的,但是由其产生的磁场是静止的。由于没有磁荷磁场是无源的,电流是静磁学中磁场唯一的旋度贡献。注意,恒 ...
静电学
静电学
电荷
库仑定律
电场
高斯定理
电势
物质中的电场
导体中的电场
导体的电容
静电能
电介质中的电场
偶极子模型
极化强度和电极化率
有电介质的高斯定理
有电介质的静电能
静电学总结
附录
\newcommand{\oiint}{\,\bigcirc\kern{-13.5pt}\int\kern{-7.2pt}\int}静电学电荷静电学的一切都要从电荷说起。实验表明自然界中只存在两种电荷——正电荷和负电荷。同种电荷相互排斥,异种电荷相互吸引。
电荷的单位是库仑(C),国际单位制中规定元电荷的大小为 $e=1.602\times 10^{-19}\text{C}$,它是由密立根通过油滴实验测得的。
注意,电荷是一种属性,而不是像电子和质子那般的实体。自然界中的物体都带有大量的电荷,不过通常由于正负电荷的数量相等,所以物体整体上是中性的。物体承载大量电荷,如果电荷能够自由移动,称为导体;如果电荷不能自由移动,称为绝缘体。
大量实验表明,在与外界没有电荷交换的系统内,正负电荷的代数和保持不变,称为电荷守恒定律。
库仑定律电荷因为相互作用而可以被观察,其相互作用 ...

分子光学
“介质对光的吸收、色散与散射,均属于分子尺度上光与物质的相互作用,故三者曾并称为分子光学。”——《现代光学基础》
光的吸收在隐逝波的讨论中,我们已经发现了线性吸收规律。从其第一性原理出发:
dI = -\alpha I dx可以推出:
I = I_0 e^{-\alpha x}其中 $\alpha$ 是吸收系数,$I_0$ 是入射光强度,$I$ 是透射光强度。
线性吸收律本质上是基于低强度光学中的线性近似。
考虑振幅表示:
\widetilde{E}(x,t) = A(x)e^{i(-\omega t + kx)}=A_0 e^{-\frac{\alpha}{2} x} e^{i(-\omega t + kx)}=A_0e^{i(-\omega t+\widetilde{k}x)}其中 $\widetilde{k} = k +i \dfrac{\alpha}{2}$,用折射率和光速表示:
\widetilde{k} = \frac{\omega n}{c} + i \frac{\alpha}{2}=\frac{\omega \widetilde{n}}{c}其中 $\widetil ...
动力学模型
分子马达
单态模型
扩散方程
速率与外力的关系
速率与ATP浓度的关系
双态模型
乌贼的巨型轴突:生物电
能斯特电势
门开关模型
去极化
带门开关的去极化模型
电报方程
钠离子通道失活
失活蛋白的回正
以下介绍动力学(含时)模型。
分子马达分子马达是细胞内负责运输和运动的蛋白质机器。它们通过化学能(通常来自ATP水解)转换为机械能,推动细胞内的物质运输。
分子马达分为以下类型:
线动马达:如肌球蛋白,沿微丝(肌动蛋白)移动,主要负责细胞收缩和物质运输。
转动马达:如噬菌体包装DNA的马达和细菌鞭毛的马达。
聚合马达
易位马达
单态模型描述分子马达需要区分内部态和外部态,其中内部态指的是马达的构象状态(如活动臂的位置和马达头部和不同分子的结合态),外部态指的是马达的具体位置。
扩散方程仅考虑马达的外部态,假设其在外力F的作用下分别以速率$k_+(F)$和$k_-(F)$前后运动,其位置概率$p(n,t)$的演化方程为:
p(n,t+\Delta t) = p(n-1,t)k_+\Delta t + p(n+1,t)k_-\Delta t+p(n,t)(1-k_-\De ...

生物网络
基因调控网络-乳糖操纵子
激活蛋白
阻遏蛋白
正负调控机制的结合
基因调控网络-果蝇胚胎
单态模型
二态模型
基因开关
生物网络是描述生物系统中各种分子、细胞或组织之间相互作用的复杂系统模型。它们通过节点(如分子、基因、蛋白质)和边(相互作用关系)来揭示生命活动的组织原则。一般使用图结构来表示。
基因调控网络-乳糖操纵子基因通过DNA转录成RNA,然后翻译成蛋白质来表达,其中起重要作用的步骤是RNA聚合酶与操纵基因的结合。存在两种调控机制:
阻遏蛋白结合在启动子上,阻止RNA聚合酶结合。
激活蛋白促进RNA聚合酶与启动子结合。
考虑聚合酶的配分函数$Z(P;N_{NS})$,其中P是聚合酶的数目,$N_{NS}$是非特异位点的数目。假设聚合酶结合在启动子上,能量为$\epsilon_{pd}^S$,结合在非特异位点上,能量为$\epsilon_{pd}^{NS}$。假设最多结合一个位点,则配分函数可以表示为:
Z(P;N_{NS}) = Z_{NS}(P,N_{NS})+Z_{NS}(P-1,N_{NS})\exp{(-\beta\epsilon_{pd}^S)}其中 ...

基础的数理化知识
化学反应式
化学平衡
非平衡动力学
郎之万方程
涨落-耗散定理
过阻尼郎之万方程及其不变分布
Kramers理论与稀有事件
伞形采样
化学反应式化学反应式是化学反应的简化表示,通常包括反应物、产物和反应条件。它们可以用来描述化学反应的平衡状态、速率和热力学性质:
A+B\rightleftharpoons C+D其中:
$A$和$B$是反应物
$C$和$D$是产物
$\rightleftharpoons$表示反应是可逆的,$\rightarrow$表示反应是不可逆的
化学平衡化学平衡是指在一个封闭系统中,反应物和产物的浓度不再随时间变化的状态。化学平衡可以用平衡常数$K$来表示:
K=\frac{[C][D]}{[A][B]}其中:
$[C]$、$[D]$、$[A]$和$[B]$分别是反应物和产物的浓度
$K$是平衡常数,表示在平衡状态下产物和反应物的浓度比值
非平衡动力学当反应不处于平衡状态时,反应速率可以用速率常数$k$来表示。以这条化学反应为例:
A~\underset{k_-}{\overset{k_+}{\rightleftharpoons}}~ B\ov ...
平衡态模型
模式系统
血红蛋白:受体配体结合模型
简单结合
多结合点位
考虑中间态
考虑协同效应
MWC模型:考虑二态
噬菌体:DNA折叠
弹性梁理论
拉伸曲线
小力情形
大力情形
插值
DNA成环自由能
细胞膜与细胞器膜
曲面的曲率
脂质双分子的自由能
膜的刚度测量
膜管
模式系统选择通用简易标准化的生物观察对象进行一类模式系统的研究,是生物学中的研究范式,其优点主要包括:
实验室可培养性
可观察及测量性
安全性
方便或针对特殊需求
按照生物结构,可以分为不同的研究层次:
结构生物学:研究大分子的三维结构及其功能关系
分子生物学与生物化学:研究大分子的功能、调控机制和动态过程
细胞生物学
遗传学
生态学
本专栏主要会设计以下模式系统:
血红蛋白:研究受体配体结合关系
噬菌体:分子马达、DNA折叠
乌贼的巨型轴突:生物电
斑马鱼的脑血管:生物运输网络
乳糖操纵子和果蝇胚胎:生物基因网络
以下展示平衡态(不含时间)的模式系统。
血红蛋白:受体配体结合模型简单结合假设体积为$\Omega$单位的溶液中有$L$个配体和1个受体,每个配体占据单位为1的体积,根据统计力学,受 ...
随机微分方程
随机微积分伊藤积分以维纳过程为基础的噪音表示为:
\xi(t) = \frac{dW(t)}{dt}显然,其具有以下性质:
均值:\langle \xi(t) \rangle = 0
方差:\langle \xi(t) \xi(t') \rangle = \sigma^2\delta(t - t')
考察一个带有噪音的随机微分方程:
\frac{dx(t)}{dt} = f(x(t), t) + g(x(t), t) \xi(t)其形式解为:
x(t) = x(0) + \int_0^t f(x(t'), t') dt' + \int_0^t g(x(t'), t') dW(t')然而,不同于黎曼积分,随机积分并不能被简单定义。由于黎曼积分的良好性质,其极限定义不依赖于积分区间的中点,即:
\int_a^b f(x) dx = \lim_{\Delta t \to 0} \sum_{i=1}^{\tau/\Delta t} f(x_i^*) \Delta t中的$x_i^*$是任意选取的点。
一般有如下两种方式来定义随机微积分:
\begin{cases}
x_i^*=x ...