动力学模型
以下介绍动力学(含时)模型。
分子马达
分子马达是细胞内负责运输和运动的蛋白质机器。它们通过化学能(通常来自ATP水解)转换为机械能,推动细胞内的物质运输。
分子马达分为以下类型:
- 线动马达:如肌球蛋白,沿微丝(肌动蛋白)移动,主要负责细胞收缩和物质运输。
- 转动马达:如噬菌体包装DNA的马达和细菌鞭毛的马达。
- 聚合马达
- 易位马达
单态模型
描述分子马达需要区分内部态和外部态,其中内部态指的是马达的构象状态(如活动臂的位置和马达头部和不同分子的结合态),外部态指的是马达的具体位置。
扩散方程
仅考虑马达的外部态,假设其在外力F的作用下分别以速率$k_+(F)$和$k_-(F)$前后运动,其位置概率$p(n,t)$的演化方程为:
移项得到差分格式:
即:
将位置概率写为连续变量$p(x,t)$,泰勒展开:
代入差分方程:
这是一个有外势场的扩散方程:
其中$D = \frac{(k_+ + k_-) a^2}{2}$是扩散系数,$v = (k_+ - k_-) a$是漂移速度。
对于平衡态,马达的速率满足详细平衡条件:
因此:
其中,$\Delta G= G(n) - G(n-1)$。有外力的时候,$\Delta G’= G(n) - G(n-1) + F a$,因此:
速率与外力的关系
现在我们只知道$k_+(F)$和$k_-(F)$的比值,假设外力只影响前向速率$k_+(F)$,而后向速率$k_-(F)$不受影响,则有:
则步进速率:
反之,如果外力只影响后向速率$k_-(F)$,而前向速率$k_+(F)$不受影响,则有:
通过实验得到,外力其实影响后向速率$k_-(F)$。
速率与ATP浓度的关系
考虑A个ATP、D个ADP和P个磷酸根在$\Omega$个格子中,对于n态,其权重为:
其中$\epsilon_{bound}$是ATP结合能。对于n+1态,其权重为:
因此,ATP水解自由能的贡献为:
如果只有前进速率$k_+$受ATP浓度影响,而后退速率$k_-(F)$不受影响,则有:
速率为:
其中,$k_+^0 = \Gamma_+ e^{-\beta \Delta G_{barrier}}/[A]$,$[A]$是标准ATP浓度。ATP水解促进向前的步进,所以$\Delta G_{hydrolysis}<0$。该模型指出了线性关系,但是ATP浓度饱和时,却没有指出速率的饱和。
如果只有后退速率$k_-(F)$受ATP浓度影响,而前进速率$k_+(F)$不受影响,则有:
速率为:
其中,$k_-^0 = \Gamma_- e^{-\beta \Delta G_{barrier}}/[A]$,$[A]$是标准ATP浓度。该模型指出了非线性关系,并且速率饱和,但是在ATP浓度较低时,速率会急剧下降。
双态模型
假设每个位点上存在两种构象状态:0和1,不同位点的跃迁即为$k_A$,同一位点的跃迁为$k_B$,则演化方程为:
记所有位点上不同状态的概率和为$P_0$和$P_1$,则有:
稳态时:
解得:
设不同状态的距离为$\delta$,不同位点的距离为$a$,速度计算为:
乌贼的巨型轴突:生物电
能斯特电势
细胞内的大分子主要带负电,如果膜对任何离子都没有通透性,那么膜两边的电位差为0。实际上,内部固定的负电荷会吸引正离子,使得膜内外的离子分布不均匀,形成电位差。
根据玻尔兹曼分布,在膜内外寻找到相应离子的概率为:
则浓度比为:
即可得到能斯特方程:
其中$F=N_Ae$是法拉第常数。
由于膜主要对$K^+$通透,因此能斯特电势主要由$K^+$决定。常见离子的膜内外浓度及其能斯特电势如下:
| 离子 | 内部浓度 (mM) | 外部浓度 (mM) | 能斯特电势 (mV) |
|---|---|---|---|
| $K^+$ | 155 | 4 | -98 |
| $Na^+$ | 12 | 145 | +67 |
| $Cl^-$ | 4 | 120 | -90 |
| $Ca^{2+}$ | $10^{-4}$ | 1.5 | +130 |
因此,膜的静息电位和接近于$K^+$的能斯特电势,约为-90mV。注意到$\frac{RT}{F} \approx 25.7$ mV,因此热运动导致的电势差在膜电位中起到了重要作用。
门开关模型
膜通道蛋白的开关和膜内外电势差有关。考虑一个二态模型,其处于开启状态的概率和此前推导的蛋白质双态模型一样:
其中$\Delta G $是开启和关闭状态的自由能差。由实验可知,膜通道在内外电势差极大的时候喜好关闭,我们可以用偶极子在电场中的能量来类比:
其中,$\Delta G_{conf}$是构象能,$p$是偶极矩,$V_{mem}$是膜电位,$d$是膜的厚度。记:
则有:
去极化
去极化是指膜电位从静息电位(通常为负值)变为更接近零或正值的过程,从而产生动作电位。去极化通常是由于膜通道的开放,允许正离子(如$Na^+$)进入细胞,导致膜内外电位差减小。
每次去极化的过程中转移的电荷对胞内离子浓度的扰动非常小。根据平行板电容器的公式:
可以计算出每次去极化的电荷量:
与此同时,细胞内离子的浓度为:
去极化的电荷转移形成跨膜离子流,其正比于两端的化学势差和膜的电导率:
然而实际上的电流并非与电压线性相关,这启发我们固定与电导的线性关系,转而研究电导本身和门开关模型的关系。假设电导和通道开启的概率成正比,在超过阈值电压$V^*$时产生非线性跃变,即可解释膜电流的非线性特性。
带门开关的去极化模型
膜电位的微分方程为:
可得稳定解:
我们作出以下假设:$g_K$对膜电位不敏感(其响应时间为5ms),而$g_{Na}$对膜电位敏感(其响应时间为1ms):
则有:
- 当$V_{mem} < V^*$时,$g_{Na}$接近0,膜电位接近$V^K_{Nernst}$;
- 当$V_{mem} > V^*$时,$g_{Na}$接近$g_{Na}^{open}$,膜电位接近$V^{Na}_{Nernst}$。
电报方程
由数学物理方程中的典型波动方程可知,电报方程是描述电信号传播的偏微分方程:
细胞电路中不存在电感$L$,因此电报方程简化为:
其中,$RC$的单位是$[\Omega] [F]/[m]^2=[s]/[m]^2$,重新改写为以下膜电位传导的形式:
其中,$\lambda$是膜电位的特征传播长度,$\tau$是膜电位的特征衰减时间。
通过数值求解,可以解出类似波传导的形式,且发现$v\propto \sqrt{d}$,这解释为什么乌贼的神经轴突很粗。
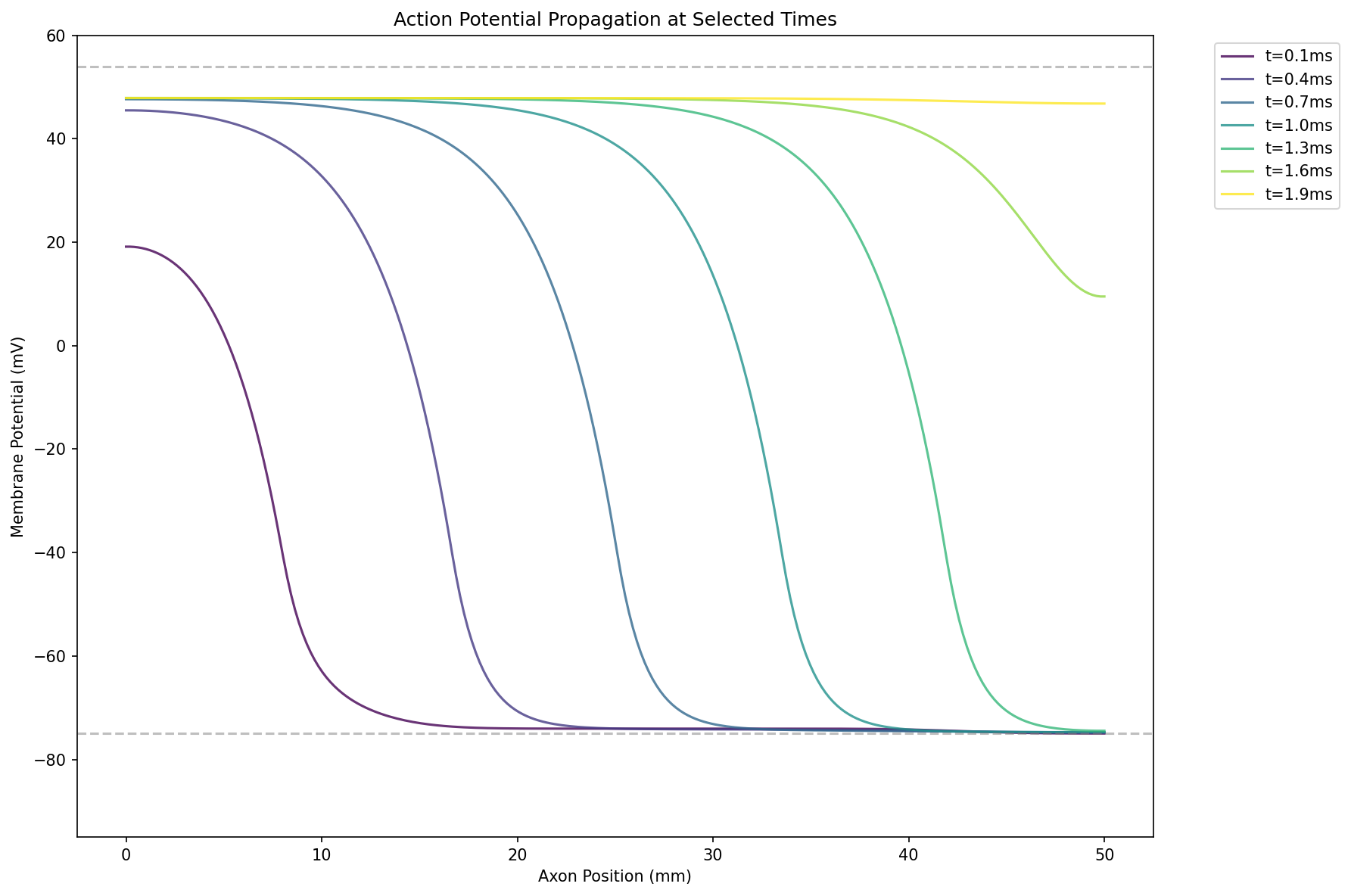
如果刺激很小,则膜电位近似指数衰减,不会产生动作电位:
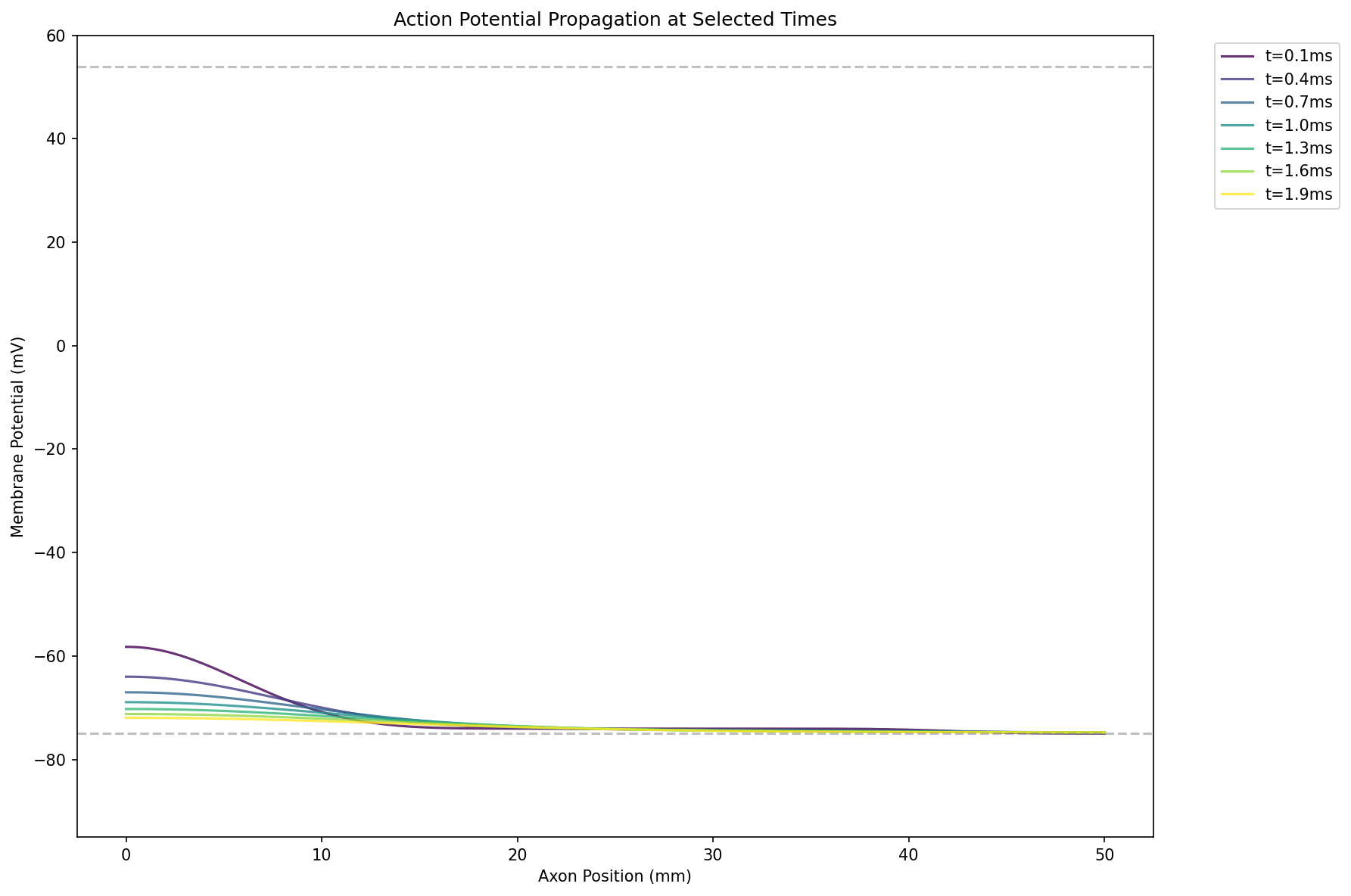
然而,目前的模型解出的波函数不能回到静息电位。这启发我们除了开启和关闭通道外,还需要考虑通道的失活。失活是指通道在开启后不久就会自动关闭,导致无法持续导电。
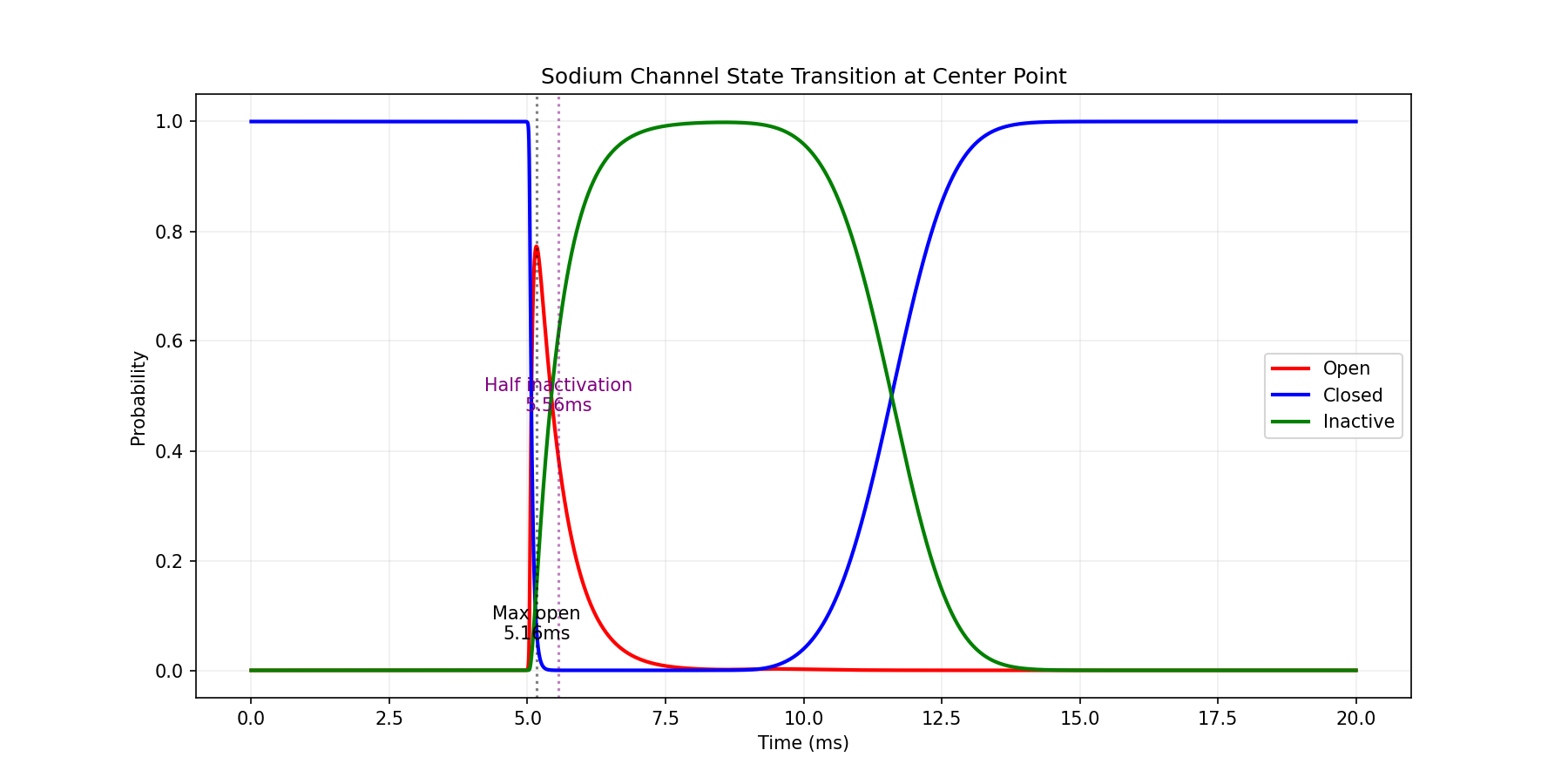
钠离子通道失活
假设存在以下状态间的转换:
进行以下简化:
重新定义$g_{Na}$:
乍一看,这和上述模型相同,不过,现在的$p_{open}$是一个三态模型的概率:
其中,通道开启速率与膜电位有关:
这就做到了动作电位产生后,通道会迅速失活,导致膜电位回到静息状态。
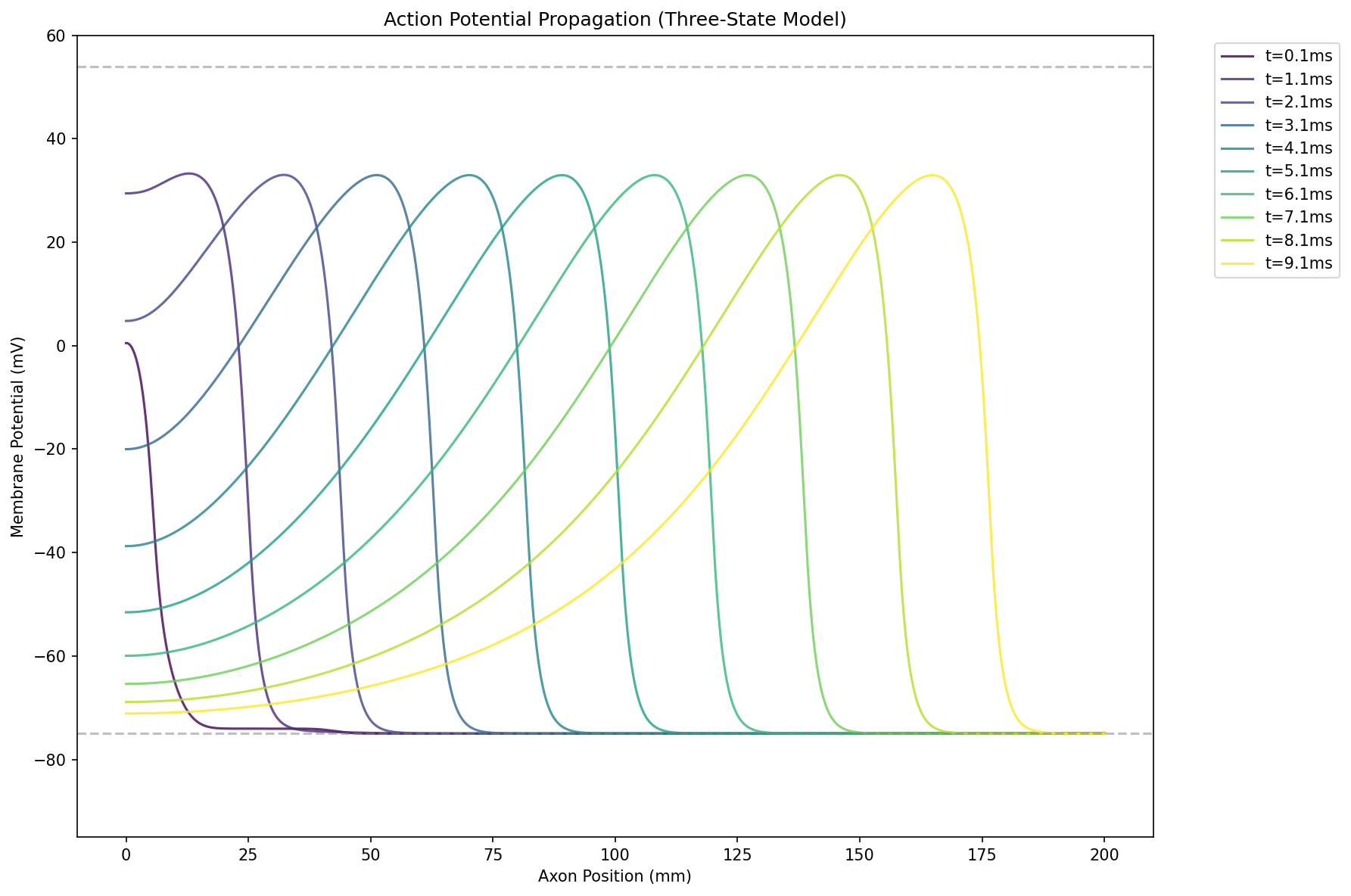
失活蛋白的回正
在原来的基础上作出以上修正:
即可得到失活蛋白向关闭蛋白的转变(这里面需要类似门开关的机制):