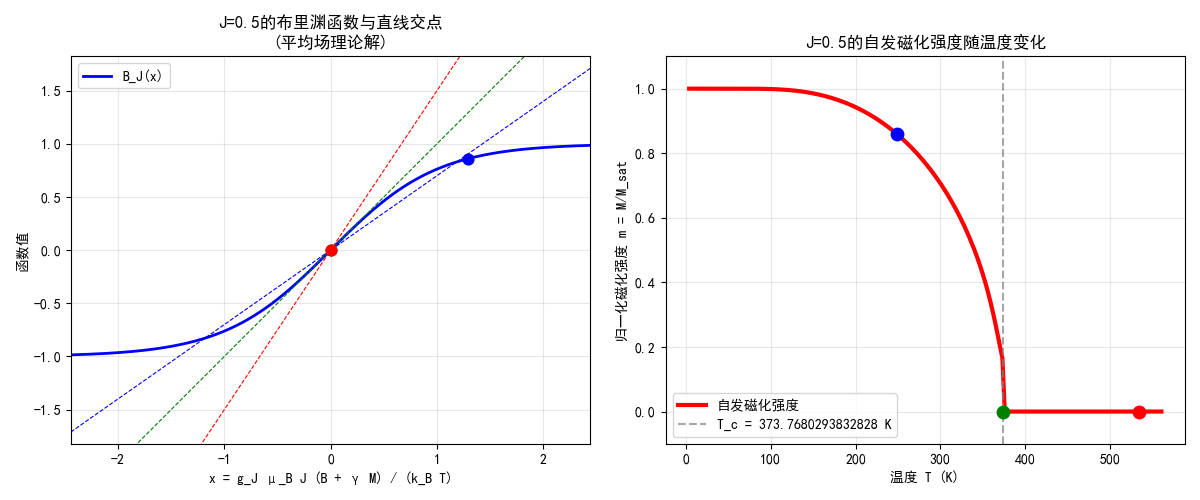
原子物理Content
作为学习量子力学的唯像桥梁,原子物理主打一个从实验中发现规律。不同于力学、电磁学、热学、光学等学科的悠长历史,原子物理学随着近代物理的发展而发展。
原子的结构
原子的精细结构
多电子原子

面试真题
普物
为什么光子没有自旋为0的态;
估算山的高度;
估算大气水蒸气凝结后有多深;
解释相速度和群速度。
量子力学
描述LS耦合的经典图像;
赝矢量和轴矢量的区别;
量子力学中有哪些势阱或势场;
画出量子谐振子基态和第一激发态的波函数图像;
密度矩阵随时间的变化;
谐振子基态能量为什么不为0;
什么是朗道能级;
氢分子的基态怎么算;
热力学统计
写出热统的三种分布规律,并说明符号的物理意义;
费米狄拉克分布随温度、能量的变化;
固体热容在高温和低温极限时,随温度或能量的变化;
理想气体的定义;
计算电子气体的熵;
玻色爱因斯坦凝聚的条件和性质;
一级相变和二级相变?举例说明。
麦克斯韦速率分布;
吉布斯佯谬。
固体物理
绝缘体和半导体有费米面吗?绝缘体有费米能量吗?
推导高温时声子热容为定值;
猜测高温时电子热容随温度的关系,并给出解释;
什么是PN结?PN结的伏安特性曲线?
什么是库珀对?产生条件和基本性质?超导体的两条性质?
铁原子外围电子排布和能级和磁矩;
解释金属、半金属、半导体、绝缘体、拓扑绝缘体的能带结构和导电性质之间的关系;
为什么电子未填满的能带可以导电,为什么Mg ...

刚体力学
刚体运动学
刚体的自由度
刚体的转动描述
刚体的转动速度描述
刚体的转动惯量
刚体的动能
刚体动力学
欧拉动力学方程
刚体运动学刚体的自由度刚体指的是系统内各质点间距离保持不变的物体。这是一种完整约束,因而可以参与自由度的减少。常见的运动对应的自由度如下:
运动类型
平动自由度
旋转自由度
总自由度
定轴转动
0
1
1
定点转动
0
3
3
平动
3
0
3
平面平行运动
2
1
3
刚体的转动描述描述刚体的平动是简单的,只需要三个质心坐标$(x_c, y_c, z_c)$即可。描述刚体的转动是复杂的,一种被动的观点是描述刚体坐标架的转动矩阵$\mathbf{U}$:
\begin{pmatrix}
i'\\
j'\\
k'
\end{pmatrix} = \mathbf{U} \begin{pmatrix}
i\\
j\\
k
\end{pmatrix}这是一个实正交矩阵。另一种主动观点为描述转动矢量或张量,而坐标架不动。
显然,刚体的位置和转动的次序是相关的,即矩阵乘法不能交换次序:
\mathbf{U}_1 \mathbf{U}_ ...

小振动
理论框架矩阵方程我们接下来讨论保守力学系统在稳定平衡位形附近作小振动的情况。解析写出拉格朗日量:
L=T-V=\sum_{i,j=1}^n \frac{m_{ij}}{2} \dot{x}_i\dot{x}_j- V(x_1, x_2, \cdots, x_n)在平衡位置附近,势能可以用泰勒公式展开:
\begin{aligned}
V(x_1, x_2, \cdots, x_n) &= V(x_{10}, x_{20}, \cdots, x_{n0}) \\&+ \sum_{i=1}^n \left( \frac{\partial V}{\partial x_i} \right)_0(x_i - x_{i0}) \\
&+ \frac{1}{2} \sum_{i,j=1}^n \left( \frac{\partial^2 V}{\partial x_i \partial x_j} \right)_0 (x_i - x_{i0})(x_j - x_{j0})\\
&+ \cdots
\end{aligned}平衡位置处$\left( \dfrac{\partial V}{\parti ...
有心力和散射问题
有心力质点所受力的作用线如果始终通过一个定点,那么称这种力为有心力。这个定点称为力心。显然,如果该力是一个保守力,则对应的势场是各向同性的。
两体问题的简化对于两个质点通过相互作用力约束在一起的问题,通过柯尼希定理,可知:
T=T_{c}+T_{r}其中 $T_{c}$ 是质点的质心动能,$T_{r}$ 是质点相对于质心的动能。
通过质点系动能定理,可知$T_c$是一个常数。展开$T_r$,有:
\begin{aligned}
T_{r} &= \frac{1}{2} m_{1} v_1^2 + \frac{1}{2} m_{2} v_2^2 \\
&= \frac{1}{2} m_{1} \left(\frac{m_2}{m_1+m_2}v\right)^2 + \frac{1}{2} m_{2}\left(\frac{m_1}{m_1+m_2}v\right)^2 \\
&=\frac12 \frac{m_1m_2}{m_1+m_2}v^2\\
&=\frac{1}{2} \mu v^2
\end{aligned}其中,$v$ 是相对速度,$\mu$ 是约化质量。这样,我们就把两体 ...
激光原理
激光(Laser)的全名是受激光辐射放大(Light Amplification by Stimulated Emission of Radiation),是通过受激辐射过程产生的相干光源。激光具有高度的单色性、方向性和相干性。
跃迁理论光的发射和吸收是光场与物质作用的结果。光场与物质作用使得组成物质的原子 (或分子)从一个本征态 (能级) 跃迁到另一个本征态,与此伴随着光场某一波型内增加或减少一个光子,这就是单光子的发射和吸收行为,这种行为包括三种基本过程,即自发辐射、受激辐射和受激吸收。
自发辐射原子中处于高能级$E_2$的电子自发地向低能级$E_1$跃迁,并发射出一个能量为$h\nu=E_2-E_1$的光子的现象,叫自发辐射。
自发辐射的速度可以用以下公式描述:
\frac{dN_2}{dt}|_{sp}=-A_{21}N_2对于没有外界扰动的情况下,可以描述为:
N_2(t)=N_2(0)e^{-A_{21}t}平均寿命为:
\tau_{21}=\frac{1}{A_{21}}常见的自发辐射的例子包括荧光和磷光。前者的寿命大概在纳秒级别,而后者的寿命由于存在自旋禁阻,大概在秒量 ...
Bloch电子动力学
Bloch电子动力学
Bloch电子的速度
半经典模型
恒定电场磁场中Bloch电子的运动
恒定电场
恒定磁场
霍尔效应
单能带模型
双能带模型
在能带论的基础上,我们不再满足经典的Drude模型和Sommerfeld模型,而是希望基于能带论建立半经典模型。
Bloch电子动力学Bloch电子的速度我们此前已经证明,周期性势场中的单电子薛定谔方程的本征函数可以写成Bloch波形式:
\psi_k^n(\vec{r})=e^{i\vec{k}\cdot\vec{r}}u_k^n(\vec{r})其中,$u_k^n(\vec{r})$是一个周期函数,满足$u_k^n(\vec{r}+\vec{R})=u_k^n(\vec{r})$,$\vec{R}$为晶格矢量。总的来说,Bloch波是由$u_k^n(\vec{r})$调幅的平面波。
将Bloch波代入薛定谔方程,得到:
\begin{aligned}
&\left[-\frac{\hbar^2}{2m}\nabla^2+V(\vec{r})\right]e^{i\vec{k}\cdot\vec{r}}u_k^n(\vec{ ...
电磁波的辐射
势和场
推迟势
李纳-维谢尔势
电偶极子的辐射
拉莫尔公式
势和场\begin{align}
\nabla\cdot\vec{E}&=\dfrac{\rho}{\varepsilon_0}\\
\nabla\cdot\vec{B}&=0\\
\nabla\times\vec{E}&=-\dfrac{\partial \vec{B}}{\partial t}\\
\nabla\times\vec{B}&=\mu_0 \vec{J}+\mu_0\varepsilon_0\dfrac{\partial \vec{E}}{\partial t}
\end{align}引入矢势$\vec{A}$和标势$V$:
E=-\nabla V-\dfrac{\partial \vec{A}}{\partial t},\quad B=\nabla\times\vec{A}这两个定义自动满足式(2)和(3)。他们需要满足的条件可以从式(1)和(4)中得到:
\begin{align}
\nabla^2V+\frac{\partial}{\partial t}\left(\nabla\cdot\vec{A} ...
电磁波
从麦克斯韦方程组到波动方程\begin{align}
\nabla\cdot\vec{E}&=\dfrac{\rho}{\varepsilon_0}\\
\nabla\cdot\vec{B}&=0\\
\nabla\times\vec{E}&=-\dfrac{\partial \vec{B}}{\partial t}\\
\nabla\times\vec{B}&=\mu_0 \vec{J}+\mu_0\varepsilon_0\dfrac{\partial \vec{E}}{\partial t}
\end{align}对式(3)取旋度:
\begin{aligned}
\text{Left}&=\nabla\times(\nabla\times\vec{E})\\
&=\nabla(\nabla\cdot\vec{E})-\nabla^2\vec{E}\\
&=-\nabla^2\vec{E}\\
\text{Right}&=\nabla\times\left(-\dfrac{\partial \vec{B}}{\partial t}\right)\\
&=-\dfrac{\part ...